3.5
Katabolické procesy
Aby mohly proběhnout v mikrobiální buňce všechny biochemické procesy, musí mít dostatečné množství energie. Z potravinářského a biotechnologického hlediska jsou nejdůležitější katabolické procesy chemoorganotrofních mikroorganismů, které tedy získávají energii oxidací organických sloučenin. Tyto oxidace katalyzují oxidoredukční enzymy. Jejich kofaktor slouží k přenosu vodíku nebo elektronů a tak se při oxidaci substrátu redukuje. Mezi koenzymy oxidoredukčních enzymů patří nikotinamidadenindinukleotid (NAD+) a jeho fosfát (NADP+), jejichž redukované formy se značí NADH a NADPH. K prostetickým skupinám těchto enzymů patří flavinové kofaktory; FAD (flavinadenindinukleotid) a FMN (flavinmononukleotid).
Při oxidaci je donorem vodíku organická látka, která se dehydrogenuje a vodíkové ionty se přenáší na akceptor, který se redukuje. Akceptorem vodíku jsou kofaktory, které přechází na svou redukovanou formu a musí být dehydrogenovány, aby mohly přenášet vodík z další molekuly substrátu. Tento proces dehydrogenace kofaktoru se nazývá regenerace kofaktoru a probíhá buď anaerobně, tedy bez účasti kyslíku nebo aerobně za spoluúčasti kyslíku.
Příklad
Při anaerobních procesech regenerace kofaktoru probíhá tak, že redukovaný kofaktor předá dva vodíky další části substrátu, takže vzniká redukovaná látka, která je ještě poměrně bohatá na energii. Mikroorganismy tuto látku nemohou dále anaerobně využít, a proto odchází z buňky jako produkt anaerobního metabolismu.
Při aerobním katabolismu jsou redukované kofaktory oxidovány za postupné účasti jednotlivých přenašečů vodíku a elektronů, tvořících tzv. dýchací řetězec. Dva vodíky redukovaných kofaktorů jsou na konci tohoto řetězce oxidovány za účasti vzdušného kyslíku na vodu. Při tomto aerobním procesu vzniká velké množství energie, která se ukládá do ATP aerobní čili oxidační fosforylací. Získávání energie aerobní fosforylací je typické pro buňky rostlinné i živočišné. ATP vzniká i při anaerobních procesech, ale v mnohem menším množství. Oxidace substrátů tedy prochází mnoha mezistupni a energie se získává postupně z některých stupňů metabolických řetězců. Kdyby se substrát oxidoval během jediné reakce, uvolnilo by se tolik tepla, že by došlo k poškození buňky (např. přímé sloučení kyslíku a vodíku je explozivní reakcí).
3.5.1
Anaerobní katabolické procesy
Souhrn
Jsou to procesy, při nichž mikroorganismy získávají energii za nepřístupu vzduchu. Někdy jim říkáme anaerobní glykolýza. Výraz je odvozen z řeckých slov: glykos = sladký a lysis = rozpouštění, rozklad. Doslovně rozpouštění cukru, česky kvašení. Mikroorganismy, které mají schopnost využívat cukry jako zdroj energie, označujeme sacharolytické. Pod správný pojem kvašení zahrnujeme rozkladné procesy bezdusíkatých látek, vyvolané enzymatickou činností saprofytických mikroorganismů.
Kvašení (fermentace) je metabolický proces, při kterém probíhá fosforylace ATP na substrátové úrovni. Z fosforylovaných meziproduktů kvašení se přenáší makroergická fosfátová skupina přímo na ADP za tvorby ATP. Jde vlastně o přenos vodíku při oxidačně redukčních procesech, o které se starají NAD-dehydrogenázy. Konečným příjemcem vodíku a elektronů je organická látka. Z energetického hlediska je kvašení proces, při němž vzniká velmi málo energie. Substrátem při fermentaci jednoduchých organických látek jsou nejčastěji glycidy. Přeměna glycidů může probíhat v několika metabolických drahách. Vždy, ale musí být hlavním meziproduktem kyselina pyrohroznová (pyruvát). Podle charakteru konečných produktů, které vznikají přeměnou kyseliny pyrohroznové a také v závislosti na původci a podmínkách celého kvasného procesu rozlišujeme několik typů kvašení.
- Etanolové kvašení
- Mléčné kvašení
- Propionové kvašení
- Máselné kvašení
- Smíšené kvašení
- Kvašení polysacharidů
- Kvašení aminokyselin
3.5.1.1
Etanolové kvašení
Patří mezi nejznámější fermentační proces využívaný při výrobě například vína, piva, droždí, krmných směsí, využívá se při kynutí těst, výrobě alkoholických mléčných nápojů atd. Zjednodušeně etanolové kvašení probíhá několika pochody, při kterých je glukóza rozkládána až na pyruvát. Pyruvát je pak anaerobně dekarboxylován na acetaldehyd a ten je redukován na etanol.
Příklad
Nejčastěji takto kvašení probíhá u kvasinek například rodu Saccharomyces, kde navazuje na glykolýzu. U bakterií například Sarcina ventriculi, Pseudomonas aj. zase probíhá drahou Entnerovou – Doudoroffovou, kde jsou konečnými produkty také etanol a oxid uhličitý. Z plísní dokáže realizovat etanolové kvašení druh Mucor javanicus.
Kvasným substrátem jsou hexózy, disacharidy, ale i polysacharidy. Kvasinky snášejí vysoké procento alkoholu 10 až 18 %. Ze substrátu spotřebují malé množství cukru k různým životním procesům, asi 2 %. Ostatní cukr zkvašují. Etanol chrání substrát před nežádoucími mikroorganismy. U etanolového kvašení není využíván kyslík. Přítomností kyslíku by byla potlačena tvorba etanolu a vznikal by hlavně oxid uhličitý a voda.
+

Obr. 14. Etanolové kvašení u kvasinek
3.5.1.2
Mléčné kvašení
Souhrn
Mléčné kvašení umožňují BMK (bakterie mléčného kvašení) rodu Lactobacillus a některé mléčné streptokoky. Hlavní funkcí těchto bakterií je rozklad cukru na kyselinu mléčnou. Kyselina mléčná chrání substrát před hnilobnými bakteriemi. Nejvíce je mléčné kvašení využíváno v mlékárenském, masném a konzervárenském průmyslu, ale také v zemědělství k výrobě siláží.
Mléčné kvašení můžeme rozdělit:
- Homofermentativní
- Heterofermentativní
Homofermentativní mléčné kvašení
U tohoto kvašení je konečným a jediným hlavním produktem kyselina mléčná. Jako substrát je zde využíván cukr, přesněji hexózy a jejich kvašení probíhá stejně jako etanolové kvašení až po tvorbu kyseliny pyrohroznové (pyruvátu). Pyruvát se následně redukuje na kyselinu mléčnou nazývanou laktát. Enzym NAD-laktátdehydrogenáza přenáší vodík na pyruvát. Připomeňme si, že BMK se vyznačují hlavně tvorbou kyseliny mléčné, nesporulují, jsou nepohyblivé. Vytváří grampozitivní tyčinky nebo koky, fakultativně anaerobní, netvoří katalázu. Jsou velice náročné na živiny a růstové faktory.
Příklad
Hlavního významu mléčného kvašení tedy inhibování růstu hnilobných bakterií se využívá k prodloužení trvanlivosti potravin (kysané zelí, kvašené okurky a zelenina). Dále se využívá při výrobě mléčných výrobků (jogurtů, tvarohů, sýrů). Mezi nejdůležitější BMK této skupiny patří Lactobacillus delbrueckii subsp. delbrueckii, Lactobacillus delbrueckii subsp. bulgaricus, Lactobacillus delbrueckii subsp. lactis, Lactobacillus helveticus a Lactobacillus acidophilus.
+
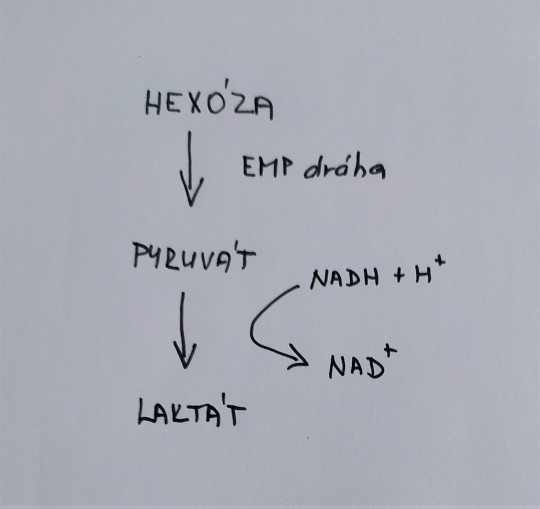
Obr. 15. Homofermentativní mléčné kvašení
Heterofermentativní mléčné kvašení
Heterofermentativní mléčné bakterie způsobují tzv. nečisté mléčné kvašení. Vyznačují se tím, že kromě kyseliny mléčné vytvářejí další různé vedlejší produkty například kyselinu octovou, jantarovou, etanol, oxid uhličitý, glycerol a vodík. Podle podmínek kvašení vzniká různé množství těchto produktů. Průměrně 40 % kyseliny mléčné, 20 % kyseliny jantarové, 10 % kyseliny octové, 10 % etanolu a zbytek bývají plyny.
Příklad
S heterofermentativním mléčným kvašením se můžeme setkat při výrobě kefíru. Často je, ale toto kvašení nežádoucí a to hlavně při výrobě ovocných šťáv a vína. Substrátem heterofermentativního mléčného kvašení mohou být pentózy například ribóza. Mezi bakterie tohoto kvašení patří například Lactobacillus brevis, Lactobacillus kefir, Lactobacillus viridescens, Leuconostoc mesenteroides, koliformní bakterie. Heterofermentativně zkvašují sacharidy také některé plísně například rod Mucor a Rhizopus.
+

Obr. 16. Heterofermentativní mléčné kvašení
Téměř všechny změny způsobené mléčnými bakteriemi mají vliv na chuť, vůni a vzhled mléka. Platí to nejen u kysaných mlék, ale i při výrobě sýrů, tedy hlavně při tvorbě jejich charakteristické chuti, vůně a konzistence. Z tohoto důvodu jsou při výrobě různých druhů mléčných výrobků používané tzv. čisté mlékařské kultury. Ty mají zajistit správně vedenou fermentaci mléka z hlediska vyráběných druhů výrobků. Velký význam mají mléčné bakterie i z hlediska zdravotního. Jsou důležité pro náš zažívací trakt, neboť vytvářením kyseliny mléčné způsobují kyselé prostředí, které brání vývoji nežádoucích hnilobných bakterií.
Propionové kvašení
Propionové kvašení je uskutečňováno hlavně propionovými bakteriemi. Propionové bakterie jsou ve velké míře zastoupeny v mléce, mléčných výrobcích, sýrech, ale i v půdě. Nachází se i v zažívacím traktu lidí a zvířat. Propionové kvašení probíhá rozkladem hexóz glykolýzou až na pyruvát nebo laktát. Vede k tvorbě propionátu, který má uplatnění jako protiplísňové činidlo. Propionové bakterie jsou anaerobní a propionovým kvašením vytváří kyselinu propionovou, octovou, oxid uhličitý a vodu.
Příklad
Činnost propionových bakterií se využívá při výrobě kyseliny propionové, ale hlavně při výrobě tvrdých sýrů. Tyto bakterie v průběhu zrání tvrdých sýrů zkvašují zbylou laktózu na kyselinu mléčnou, propionovou, octovou, vodu a oxid uhličitý. Ten vytváří typická oka v sýrech ementálského typu. Za vhodných podmínek jsou některé propionové bakterie schopny tvořit vitamin B12. Typickými představiteli propionových bakterií jsou Propionibacterium freudenreichii, Propionibacterium shermanii, Propionibacterium thoenii atd.
Máselné kvašení
Máselné kvašení způsobují představitelé rodu Clostridium. Bakterie rodu Clostridium jsou striktně anaerobní. Kvašením cukrů (hexóz) vzniká kyselina máselná, silně zapáchající látka a plyny CO2, H2, kyselina octová, kapronová, kaprylová, aceton a butylalkohol. Tento proces dokáže znehodnotit potraviny, ale je také užitečný při biologickém čištění odpadních vod. Mezi hlavní představitele máselného kvašení řadíme Clostridium butyricum, Clostridium acetobutilicum. Tyto bakterie jsou hojně rozšířeny v půdě, hnoji, znečištěných vodách. Máselné kvašení je nežádoucí při konzervaci potravin a při výrobě krmiv. Kyselina máselná není jedovatá, ale odporně páchne. Zároveň znehodnocuje potraviny chuťově. Původci máselného kvašení způsobují hnilobný rozklad bílkovin a tím se potraviny stávají nepoživatelnými. Rozvoj bakterií máselného kvašení zastavíme rychlým okyselením potraviny na pH 4,2.
Smíšené kvašení
Smíšené kvašení vyvolávají bakterie z čeledi Enterobacteriaceae, jako jsou rody Escherichia, Enterobacter, Salmonella, ale i bakterie rodu Bacillus a některé druhy rodu Proteus, Pseudomonas a Serratia. Tímto kvašení vzniká mnoho různých produktů. Mezi nejčastější patří kyselina octová, mravenčí, mléčná, jantarová, ale i acetoin, 2,3-butandiol, vodík a oxid uhličitý. Kyseliny tvoří hlavně bakterie rodu Escherichia, Salmonella, Proteus. Kyselinu mléčnou, acetoin, 2,3-butandiol tvoří bakterie rodu Enterobacter, Bacillus, Serratia. Máselné aroma vzniká oxidací acetoinu na vzduchu za vzniku biacetylu, což je žádoucí hlavně při zrání smetany. Produkuje ho Leuconostoc cremoris. Naopak produkce biacetylu rodem Pediococcus, nepříznivě ovlivňuje jakost piva.
Kvašení polysacharidů
Některé bakterie mají schopnost využívat při kvašení jako substrát také polysacharidy. Pomáhají tomu exoenzymy, které štěpí tyto látky na látky jednodušší. A ty pak mohou vstupovat do buňky.
Kvašení celulózy
Celulóza je polysacharid velmi odolný proti rozkladné činnosti mikroorganismů. V přírodě se tvoří celulózy velmi mnoho. V rostlinných zbytcích dosahuje její obsah 45 až 80 %. Štěpení celulózy se účastní dva enzymy, celuláza a celobiáza, které ji hydrolyzují na glukózu. Mikroorganismy rozkládají celulózu za podmínek aerobních i anaerobních.
Poznámka
Celulóza je pro člověka nestravitelná. Dobře ji však tráví býložravci. A to proto, že v jejich trávicím traktu jsou přítomny právě celulózové bakterie. Mezi tyto bakterie patří anaerobní druhy rodu Clostridium.
Výsledným produktem zkvašování glukózy je kyselina mravenčí, octová, máselná, mléčná, etanol, oxid uhličitý a vodík. V případě přítomnosti metanových bakterií je oxid uhličitý redukován na metan.
Kvašení glykogenu a škrobů
Glykogen a škrob jsou štěpeny pomocí amylolytických enzymů, které vytváří různé bakterie. Nejznámější jsou zástupci rodu Clostridium a Bacillus. Ty vytváří α-amylázy, které svým působením rozkládají škrob a glykogen na oligosacharidy s glukózovými jednotkami a ty jsou v konečné fázi rozštěpeny na maltózu. Maltóza je za pomoci enzymu maltáza hydrolyzována na glukózu a ta zkvašována na konečné produkty.
Kvašení pektinu
K rozkladu pektinů dochází s pomocí enzymů protopektinázy. Činností těchto enzymů vzniká celulóza a protopektiny, které tvoří společně s polysacharidy komplexy. Ty jsou dále štěpeny na pektin a složky určitého komplexu. Působením enzymu pektináza je z pektinu odštěpen metylalkohol a vzniká kyselina pektinová. Hydrolýzou kyseliny pektinové za pomoci pektolázy vzniká kyselina galaktouranová, arabinóza, galaktóza a xylóza. Ty mohou být dále zkvašovány na kyselinu máselnou, octovou, oxid uhličitý a vodík. Původci mohou být například některé druhy rodu Clostridium nebo Bacillus.
Kvašení aminokyselin
Zkvašovány kromě sacharidů mohou být i dusíkaté organické látky například aminokyseliny. Aminokyseliny jsou rozkládány proteolytickými enzymy. Endopeptidázami jsou štěpeny peptidické vazby podél peptidických řetězců a exopeptidázami zase koncové články těchto řetězců. Společně je molekula bílkoviny rozštěpena na oligopeptidy a následně až na volné aminokyseliny. Ty jsou pak přenášeny do buňky a dále rozkládány. Mikroorganismy využívající bílkoviny a štěpné produkty aminokyselin označujeme proteolytické.
Rozklad bílkovin
Mikrobiálnímu rozkladu bílkovin (proteolýze) říkáme obecně hniloba. Je to enzymový, biochemický proces, při němž mikroorganismy vylučují do prostředí extracelulární enzymy, zvané proteázy, které hydrolyzují peptidové vazby a odbourávají makromolekuly bílkovin na složky o malé molekulové hmotnosti. Rozklad bílkovin probíhá v několika stupních. Bílkovina je štěpena na peptony rozpustné ve vodě, popř. polypeptidy, dále na oligopeptidy a dipeptidy, které se štěpí na aminokyseliny, amoniak a těkavé kyseliny. Proteolytické bakterie tento proces tvoří za aerobních i anaerobních podmínek. Tyto hnilobné procesy mají velký význam v přírodě, protože se tak uvolňují z odumřelých těl anorganické sloučeniny biogenních prvků, zvláště C, N, S a P. Rozkladem vznikají jednoduché sloučeniny, které jsou v této podobě nebo po další přeměně asimilovány vyššími organismy, a tak udrženy v přirozeném koloběhu prvků.
Poznámka
Proteolýza je základem postupné mineralizace organických látek. V potravinářském průmyslu způsobuje tento proces většinou vady. Výjimkou je zrání sýrů, kde je rozklad bílkovin základním procesem, pro tvorbu typické struktury, chuti a vůně.
Rozklad tuků
Mikrobiální rozklad tuků je způsoben tzv. lipolytickými mikroorganismy, které svými enzymy (lipázami) katalyzují rozklad tuků. Lipolytické mikroorganismy štěpí (hydrolyzují) tuky rozkladem esterické vazby na glycerol a mastné kyseliny. Další mikroorganismy oxidují mastné kyseliny až na CO2 a H2O. Tyto procesy mají opět velký význam při rozkladu v půdě.
Zajímavost
Je nutné zdůraznit zcela ohromující skutečnost – totiž, že všechny živé organismy, včetně mikroorganismů, metabolizují glukózu až po tvorbu pyruvátu zcela stejným mechanismem a přitom se mohou od sebe podstatně odlišovat stupněm tělesné organizace, morfologicky i velikostí.
3.5.2
Aerobní katabolické procesy
Bakterie ve většině případů získávají potřebnou energii oxidací organického substrátu molekulovým kyslíkem tedy aerobní respirací. Prostřednictvím kompletního transportního řetězce dochází k přenosu vodíku a elektronů až na kyslík, který se slučuje s vodíkem na vodu. Současně dochází k postupné oxidaci organického substrátu. Proces může probíhat jako úplná oxidace substrátu, kdy je substrát oxidován na oxid uhličitý nebo jako, neúplná oxidace substrátu, kdy je po vytvoření určitého metabolitu zastaven a dojde k hromadění.
3.5.2.1
Neúplná oxidace substrátu
Substrátem bývají alkoholy a sacharidy. Z primárních alkoholů vznikají mastné kyseliny a oxidace probíhá přes aldehyd, který je dále oxidován na kyselinu. Sekundární a terciální alkoholy bývají oxidovány za vzniku ketonů. Ze sacharidů podléhá neúplné oxidaci hlavně glukóza a vzniká kyselina glukonová. Tohoto procesu se účastní především zástupci rodu Acetobacter a Gluconobacter. Oxidace glukózy probíhá na dráze pentózového cyklu, ale může se uskutečňovat i bez tvorby fosfátů.
3.5.2.2
Úplná oxidace substrátu
Tyto procesy probíhají na metabolických drahách, které mají cyklický charakter. Jsou to Krebsův cyklus (cyklus kyseliny citrónové, citrátový), glyoxalátový cyklus a pentózový cyklus.
Krebsův cyklus
Jsou to dýchací pochody aerobní povahy, při nichž se organický substrát přeměňuje až na oxid uhličitý a vodu. Aerobní procesy poskytují buňkám daleko více energie než procesy anaerobní. Substrátem v Krebsově cyklu mohou být jak sacharidy tak třeba lipidy, bílkoviny nebo další organické sloučeniny. Substrát je postupně přeměňován na acetyl-Co-A. Odbourávání většiny substrátů končí ve druhé fázi aerobního katabolismu acetylkoenzymem A. Vlastní cyklus začíná kondenzací acetyl-Co-A s oxalacetátem za vzniku kyseliny citrónové. Postupně pak dochází k přeměnám citrátu zpět až na oxalacetát. Meziprodukty citrátového cyklu jsou trikarboxylové kyseliny citrónová a izocitrónová a také dikarboxylové kyseliny α-oxoglutarová, jantarová, fumarová, jablečná a oxaloctová. Kromě acetátu a pyruvátu jsou citrátovým cyklem využívány i konečné produkty glykolýzy jako etanol, glycerol, laktát, dikarboxylové a trikarboxylové kyseliny, vyšší mastné kyseliny a velká většina aminokyselin.
Bílkovinný substrát se v citrátovém cyklu přeměňuje rozštěpením proteolytických enzymů na aminokyseliny. A aminokyseliny jsou odbourávány buď deaminací (vzniknou amoniak a organické kyseliny, které jsou dále metabolizovány), nebo transaminací (aminové skupiny jsou přenášeny za pomoci enzymů na α-ketokyseliny za vzniku jiné aminokyseliny). Aminokyseliny jako alanin, kyselina glutamová a asparagová se po deaminaci přímo účastní citrátového cyklu jako meziprodukty a leucin, lyzin, tryptofan, metionin, valin se musí přeměnit na další meziprodukty jako je acetyl-Co-A, pyruvát a oxalacetát. Konečnými produkty oxidace aminokyselin závisí na typu bakterií a mohou to být acetyl-Co-A, kyselina pyrohroznová, acetoctová, fumarová, jantarová a další.
Lipidový substrát se pomocí enzymů lipáz štěpí na mastné kyseliny a glycerol. β-oxidací jsou mastné kyseliny dále odbourávány. K odbourávání dochází tak, že mastné kyseliny se oxidují za vzniku acetyl-Co-A a kyseliny o dva uhlíky kratší, než je původní mastná kyselina. Pak dojde k další oxidaci za opětovného zkrácení řetězce a proces se opakuje, dokud se mastná kyselina nerozloží. Rozložené acetylové zbytky jsou vázané na Co-A. AcetylCo-A, který vznikne, se může dál oxidovat v citrátovém cyklu nebo může být součástí biosyntetických dějů.
Glycerol uvolněný rozkladem lipidů se oxiduje glykolýzou přes pyruvát. Pyruvát je pak v citrátovém cyklu aerobně oxidován na vodu a oxid uhličitý.
Glukóza než vstoupí do citrátového cyklu je glykolýzou přeměněna na pyruvát a ten aerobně s pomocí enzymů na acetyl-Co-A.
Citrátový cyklus je tzv. metabolickým uzlem, ve kterém se setkávají katabolické a anabolické dráhy a proto je součástí obou. Takové dráhy nazýváme amfibolické. Krebsův cyklus slouží mikroorganismům jako zdroj meziproduktů pro syntézu aminokyselin a jiných látek. Kdyby se odčerpal, například oxalacetát pro biosyntézy během buněčného růstu nebylo by dostatek materiálu pro regeneraci oxalacetátu a celý cyklus by se zastavil. Proto jsou mikroorganismy vybaveny pomocnými cykly. Tyto cykly, tedy jejich náhradní reakce doplňují odčerpané meziprodukty a tím cyklus může pokračovat. Říkáme jim anaplerotické.
Glyoxalátový cyklus
Díky tomuto cyklu mohou mikroorganismy využívat acetát nebo mastné kyseliny ve formě acetyl-Co-A jako jediný zdroj uhlíku a energie. Dále je to způsob doplňování odčerpaných meziproduktů citrátového cyklu u bakterií bez přísunu pyruvátu. Tento cyklus byl zjištěn především u rodu Pseudomonas a Xanthomonas. Celý tento cyklus má velké množství enzymů a reakcí společných s citrátovým cyklem. Rozdíl je jen v chybějících enzymech, které štěpí izocitrát na sukcinát a glykoxalát. Někdy probíhají oba cykly současně. Enzymy jak citrátového tak glyoxalátového cyklu jsou u eukaryotních mikroorganismů v mitochondriích a u bakterií (prokaryotních mikroorganismech) v cytoplazmatické membráně.
3.5.2.3
Anaerobní respirace
Některé druhy mikroorganismů získávají energii i za anaerobních podmínek oxidačně redukčními reakcemi. V anaerobním prostředí je konečným akceptorem elektronů anorganická sloučenina. Přesněji řečeno akceptorem elektronů a vodíku je kyslík vázaný v SO42-, NO3-, CO32-, a jiné sloučeniny.
Podle povahy redukované látky rozeznáváme:
- respiraci nitrátů
- respiraci síranů
- respiraci CO2
- respiraci fumarátu
- respiraci tetrathionátu
Respirace nitrátů
Jedna z nejrozšířenějších anaerobních respirací u bakterií. Konečným akceptorem elektronů je NO3-, který je redukován na NO2-. U několika málo bakterií dokonce až na N2. Jev, kdy se uvolňuje dusík, nazýváme denitrifikací. A v obou případech nitrátové respirace jsou reakční produkty jako dusík nebo dusitany vylučovány do prostředí. Respiraci nitrátů si však nesmíme plést s asimilací nitrátu jako zdroje dusíku, kdy dochází k asimilační reakci NO3- na NH3. U respirace nitrátů jde o disimilativní redukci nitrátu.
Nitrátovou respiraci využívá mnoho bakterií, ale nikdy to pro ni není jediný způsob získávání energie.
Příklad
Redukce nitrátů na nitrity byla prokázána u rodů Escherichia, Shigella, Salmonella, Citrobacter, Serratia, Proteus, Staphylococcus, Mycobacterium, Bacillus, Lactobacillus, Clostridium, Pseudomonas, Micrococcus atd.
Redukce NO3- na N2 je uskutečňována denitrifikačními bakteriemi jako jsou například Bacillus denitrificans, Bacillus stutzeri. Denitrifikace probíhá v půdě i vodě. Celý tento proces ovlivňuje koloběh dusíku v přírodě.
Asimilativní redukce NO3- na NH3 probíhá u bakterií rodu Escherichia, Bacillus, Micrococcus, Clostridium, ale i u kvasinek rodu Candida, Rhodotorula, Hansenula a plísní rodu Aspergillus, Penicillium.
Respirace síranů
Anaerobní respiraci síranů dokáží provádět pouze dva rody bakterií a to rod Desulfovibrio a Desulfotomaculum. Důležité je také zmínit, že je to jejich hlavní způsob získávání energie. Tyto bakterie žijí v anaerobních podmínkách například v bahně a redukují sírany na sulfan. Sulfan reaguje s kovy za vzniku černých sulfidů. Dále tyto bakterie slouží jako indikátor anaerobních procesů ve vodách. Produkce sulfanu nepříznivě ovlivňuje kvalitu vody. Podílejí se také na koloběhu síry v přírodě, umožňují život fototrofním bakteriím v anaerobních podmínkách a sirným bakteriím v aerobních podmínkách.
Respirace oxidu uhličitého
Této respiraci také říkáme metanové kvašení, které způsobují jen některé bakterie striktně anaerobní. Například rody Methanobacterium, Methanococcus, Methanosarcina. Jako akceptor elektronů využívají CO2 a tím dochází k redukci na CH4. Produkce metanu je velmi intenzivní. Uskutečňuje se anaerobním rozkladem organické hmoty třeba v bahně, jezerních sedimentech a v zažívacím traktu přežvýkavců. Tuto respiraci využíváme při čištění odpadních vod, při likvidaci živočišného odpadu a při výrobě bioplynu.
Poznámka
Bakterie těchto rodů mají velký ekologický význam, protože mohou navazovat na fermentační procesy v anaerobním prostředí, kterými se nahromadí organické kyseliny anaerobně již dále nezpracovatelné. Bakterie produkující metan je mohou využít k redukci oxidu uhličitého a tím vracet uhlík zpět do koloběhu.
Respirace fumarátu
Fumarát je jako akceptor vodíku a elektronů redukován na sukcinát, který se hromadí v prostředí. Respirace fumarátu tvoří ATP na membráně oxidační fosforylací. Tuto respiraci uskutečňují bakterie rodu Escherichia, Enterobacter, Citrobacter, Shigella, Proteus, Aeromonas apod.
Respirace tetrathionátu
Tuto respiraci uskutečňují enterobakterie Salmonella, Serratia, Proteus, Citrobacter, ale ne Escherichia.
Na závěr můžeme říct, že některé bakterie mohou využívat i několik typů anaerobní respirace a to někdy i vedle aerobní respirace a fermentace.